dr inż. Katarzyna Tkacz
mgr inż. Joanna Ewelina Rydel
dr hab. inż. Monika Modzelewska-Kapituła, prof. UWM
mgr inż. Weronika Zduńczyk
Katedra Technologii i Chemii Mięsa,
Wydział Nauki o Żywności
Uniwersytet Warmińsko-Mazurski w Olsztynie
Zdolność utrzymywania wody przez mięso oraz jego przetwory jest jednym z ważniejszych aspektów z punktu widzenia technologicznego, jak i ekonomicznego. Służy jako parametr do oceny jakości mięsa oraz jest decydującym wskaźnikiem przydatności technologicznej [12]. Cecha ta ma znaczący wpływ na soczystość i kruchość mięsa oraz na zmiany zawartości wody w mięsie w trakcie transportu, chłodniczego przechowywania, a także podczas obróbki termicznej [6].

Struktura wody oraz jej interakcja z białkami
Woda to podstawowy składnik zwierzęcej, oraz roślinnej komórki biologicznej. Jest to związek reaktywny, który pełni rolę uniwersalnego rozpuszczalnika oraz czynnika dyspergującego [1; 7]. Cząsteczka wody dysponuje biegunowym przestrzennym układem ładunków elektrycznych, a więc jest dipolem, który jest przyciągany do naładowanych cząstek, takich jak białka [4]. Dwa atomy wodoru, połączone są kowalencyjnie z danym atomem tlenu, w rozmieszczeniu symetrycznym, a protony wodorów wytwarzają biegun posiadający ładunek dodatni. W atomie tlenu występują również dwie strefy kumulowania elektronów, a biegun ładunku przyjmuje wartość ujemną [5]. Cząsteczki wody są w stanie wzajemnie łączyć się poprzez wiązania wodorowe, natomiast ujemny region elektronowy poszczególnej cząsteczki zostaje przyłączony przez dodatni region następnej cząsteczki wody [7].
Przyciąganie oraz wiązanie cząsteczki wody następuje poprzez obdarowanie ładunkiem elektrycznym poszczególnych reaktywnie odmiennych związków. Rozpuszczalniki zjonizowane takie jak kwasy, zasady czy sole, posiadają jony zawierające ładunki elektryczne, które zostają wprowadzone do uporządkowanej struktury wody, gdzie zmienia się ich układ przestrzenny oraz zachodzi mocna polaryzacja dipolów wody. Cząsteczki wody są wzajemnie przyciągane oraz łączone poprzez posiadane przez nie ładunki [1].
Białka miofibrylarne stanowią 55-60% białek we włóknach mięśniowych i są ważnym składnikiem odpowiadającymi za wiązanie wody oraz utrzymanie jej w mięśniach [2]. Miofibryle stanowią większą część komórek mięśniowych, a ich zawartość to 82-87% w stosunku do całej objętości komórek mięśniowych [4]. Powiązane jest to z ich właściwościami fizykochemicznymi, a także szczególną trójwymiarową strukturą, zmianami przedubojowymi oraz zmianami, jakie zachodzą w trakcie przechowywania mięsa [1]. Znacząca ilość wody znajduje się w miofibrylach, pomiędzy przestrzeniami grubych filamentów miozynowych a cienkimi filamentami aktynowymi (w tzw. kieszeniach wodnych). Wielkość przestrzeni pomiędzy filamentami kształtuje się w zakresie od 320 do 570 A i jest uzależniona od wartości pH, siły jonowej oraz ciśnienia osmotycznego i stanu mięśnia po uboju [7; 5]. Zawartość wody związanej poprzez białka uzależnione jest od liczby grup polarnych białka, które posiadają ładunek dodatni oraz ujemny, odległości pomiędzy grupami polarnymi struktury białek oraz wielkości przestrzeni, w której mają możliwość ustabilizowania się. Cząsteczki wody w trakcie wiązania się z grupami posiadającymi ładunek oraz z grupami polarnymi białka obniżają natężenie pola elektrycznego wokoło tych grup, a także powodują stabilizację układu białko – woda poprzez zastosowanie zmniejszonej wolnej energii, jaką dany układ posiada [5; 4]. Grupy polarne umiejscowione są na powierzchni zewnętrznej cząsteczki białka i są ściśle powiązane z cząsteczkami wody. Siła z jaką przyciągane są cząsteczki przez grupy polarne, ulega zmniejszeniu wraz ze wzrostem odległości. Reszta przylegających warstw wody jest słabiej wiązana. Wymiana następuje ciągle pomiędzy cząsteczkami wody mocniej i słabiej związanej występującej na powierzchni cząsteczki białka [5].
Rozmieszczenie wody w mięsie
Najwięcej wody znajduje się w tkance mięśniowej tuszy zwierzęcej, natomiast jej zawartość w tkance tłuszczowej jest znikoma. Rozmieszczenie jej jest również zróżnicowane. Około 85% całej zawartości wody znajduje się we włóknach mięśniowych (około 60% jest w miofibrylach, natomiast około 25% występuje w sarkoplazmie), a około 15% znajduje się w przestrzeniach poza włókienkowych [6]. Ilość wody występująca w mięśniach jest różna w zależności od gatunku i rasy zwierząt. Różnice te zależą od ilości tłuszczu śródmięśniowego – im zawartość tłuszczu w mięśniu jest większa, tym zawartość wody jest mniejsza. Mięśnie, które cechowały się większą aktywnością przyżyciową, zawierają większą ilość wody, niż te mięśnie, które oznaczały się mniejszą aktywnością [5].
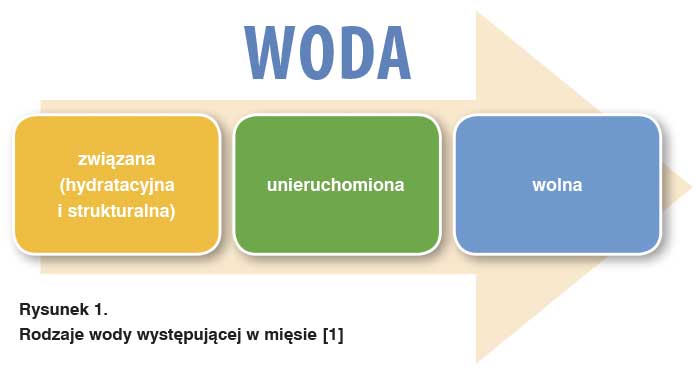
Woda, która w tkance mięśniowej jest środowiskiem, gdzie zachodzą procesy życiowe takie jak: przemiana materii oraz energii, transport substancji odżywczych oraz produktów odpadowych, odgrywa także znaczącą rolę w przemianach poubojowych. Biorąc udział w utrzymywaniu konformacji białek, a także w kształtowaniu reologicznych cech mięsa, w znaczący sposób wpływa na jakość surowca mięsnego [7; 1]. Forma występowania wody w mięsie oraz stopień hydratacji, a także związania wody są przyczynowo zróżnicowane i zależne od stanu fizycznego białek. Rodzaje wody występującej w mięsie przedstawiono na rys.1.
Woda związana
Woda związana występuje w ilości około 5% całkowitej ilości wody [5]. Jest ona wiązana poprzez grupy polarne i zjonizowane białek, a także innych niewodnych składników występujących w postaci warstw monomolekularnych. Woda jest wiązana przede wszystkim przez białka miofibryli, a także poprzez białka tkanki łącznej [7]. Ilość wody związanej zmienia się w niewielkim stopniu w trakcie dojrzewania mięsa podczas występowania stężenia pośmiertnego [5]. Zawartość w mięsie wody związanej nie jest zależna od zmian struktury mięsa, m.in. jego rozdrobnienia czy oddziaływania czynników fizykochemicznych i nie jest możliwe usunięcie jej przy zastosowaniu sił zewnętrznych tj. nacisk lub wirowanie [14].
W obrębie wody związanej możemy wyróżnić wodę hydratacyjną i strukturalną. Jeśli cząsteczki wody są unieruchomione, a ich ruch wynika jedynie z ruchu makrocząsteczek, określa się ją jako strukturalną. Gdy oddziaływania występujące w układzie woda – substancja rozpuszczona nie ograniczają całkowicie ruchu cząsteczki wody, mówimy o wodzie hydratacyjnej [15].
Woda unieruchomiona
Największą część wody stanowi woda unieruchomiona, która jest rozmieszczona w postaci pozornie wolnych cząsteczek występujących w przestrzeniach kapilarnych wewnątrzkomórkowych oraz międzykomórkowych mięśnia. Woda unieruchomiona utrzymywana jest poprzez siły elektrostatyczne, a także kapilarne [7; 4]. Największą część w całkowitej ilości wody (80%) stanowi właśnie woda unieruchomiona, która znajduje się w miofibrylach oraz w błonie komórkowej [11].
W strukturze mięśnia występuje wiele przestrzeni kapilarnych, gdzie istnieje możliwość unieruchomienia wody [7]. Są to: przestrzenie występujące pomiędzy grubymi filamentami miozynowymi oraz cienkimi filamentami aktynowymi znajdujące się w obrębie miofibryli, przestrzenie pomiędzy miofibrylami oraz pomiędzy miofibrylami i błoną komórkową, jak również przestrzenie między komórkami oraz przestrzenie pomiędzy wiązkami włókien mięśniowych [5].
W okresie przyżyciowym zwierzęcia woda niezwiązana zostaje unieruchomiona w mięśniach w przestrzeniach międzycząsteczkowych oraz wewnątrzcząsteczkowych. Czynnikiem, który odpowiada za unieruchomienie wody, jest ujemne ciśnienie hydrostatyczne, jakie panuje w okolicy przestrzeni śródmięśniowych [7; 5]. Siła, z jaką unieruchomiona jest woda, zależy przede wszystkim od przestrzeni kapilarnych: im średnica przestrzeni jest mniejsza, tym silniejsze jest unieruchomienie wody, a siła potrzebna do jej usunięcia jest większa. Znaczący udział w retencji wody posiadają przestrzenie kapilarne, które znajdują się w strukturze miofibryli, gdyż większa ilość wody jest unieruchamiana w otoczeniu cienkich filamentów aktynowych niż w otoczeniu grubych filamentów miozynowych [5; 7].
Istnieje także możliwość dyfuzyjnej wymiany cząsteczek wody z komórek w kierunku przestrzeni międzykomórkowych i na odwrót, a także możliwość parowania wody z powierzchni tkanki w kierunku atmosfery [2]. We wczesnej fazie okresu poubojowego, poprzedzającej stężenie pośmiertne, woda unieruchomiona nie wypływa naturalnie z mięsa, usunąć ją można natomiast, stosując suszenie bądź wymrożenie [7].
Przemiany, jakie zachodzą w strukturze mięśnia i właściwościach białek mięśniowych w trakcie rozwoju stężenia pośmiertnego, transformacji mięśni w mięso oraz także w trakcie obniżenia pH, znacząco wpływają na unieruchomienie wody, a także na jej rozmieszczenie w przestrzeniach kapilarnych mięśnia [5]. Powoduje to, że część wody wcześniej unieruchomionej może stać się tzw. wodą wolną i wypłynąć z mięsa w formie wycieku [7].
Woda wolna
Woda wolna wypływa z mięsa pod postacią wycieku w sposób swobodny, a przepływ z tkanki odbywa się bez zakłóceń [4; 8]. Właściwości wody wolnej są zbliżone do tych, które charakteryzują wodę w rozcieńczonych roztworach soli – cząsteczki wody są połączone wiązaniami wodorowymi i charakteryzuje je zwiększona ruchliwość. Woda wolna nie występuje w mięśniach przed stężeniem pośmiertnym.
Woda ta stanowi rozpuszczalnik dla substancji organicznych oraz nieorganicznych, a także bierze udział w procesach fizyczno-chemicznych oraz jest luźno powiązana z białkami [7]. Słabe siły powierzchniowe dają możliwość zatrzymywania wody wolnej w mięsie, a jej ilość zwiększa się w warunkach, które pozwalają na przemieszczanie się wody z przestrzeni kapilarnych ku powierzchni mięsa [5].
Po przecięciu mięsa następuje wypływanie wody wolnej, pod wpływem sił grawitacyjnych, podłużnymi kanałami pomiędzy pęczkami włókien na zewnątrz [13].
Zmiany stopnia uwodnienia mięsa
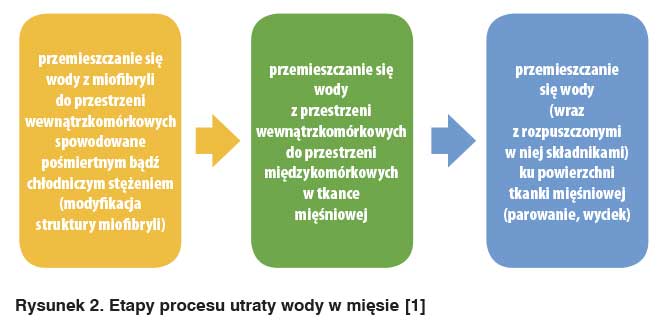
Straty wody, jakie występują w mięsie w trakcie przechowywania chłodniczego, mrożenia oraz procesów technologicznych definiuje się jako wyciek [4; 9]. Z histologicznego punktu widzenia, stratę wody oraz składników w niej zawartych, można określić jako trójstopniowy proces, którego poszczególne etapy przedstawiono na rys. 2.
Jednym z czynników mającym wpływ na zmianę stopnia uwodnienia są przemiany biochemiczne, które związane są ze stanem rigor moritis oraz dojrzewaniem mięsa, w którym zostaje zmieniona całkowita konfiguracja przestrzenna oraz ładunek elektrostatyczny białek miofibrylarnych, powodując zmianę cech jakościowych, a także właściwości funkcjonalnych białek [2].
Literatura
1. Dolatowski Z., Twarda J., 2002, Rola wody w mięsie, Mięso i Wędliny, 32-35.
2. Dolatowski Z. Stadnik J., Olszak M., 2006, Wpływ sonifikacji mięsa wołowego (m. semimembranosus) na wodochłonność, wyciek cieplny i strukturę po rozmrożeniu, Rocznik Instytutu Przemysłu Mięsnego i Tłuszczowego, 44(1), 93-101.
3. Domaradzki P., Skałecki P., Florek M., Litwińczuk Z., 2010, Związek kolagenu z wybranymi parametrami technologicznymi mięsa wieprzowego, ŻYWNOŚĆ. Nauka. Technologia. Jakość., 17(4), 50-62.
4. Huff-Lonergan Elisabeth, 2010, Water-holding capacity of fresh meat, National Board meat pork.
5. Kołczak Tadeusz., 2007, Retencja wody w mięsie, Gospodarka Mięsna, 30-35.
6. Modzelewska-Kapituła M., Cierach M., 2012, Wykorzystanie komputerowej analizy obrazu do oznaczania zawartości wody wolnej w mięsie metodą Graua- Hamma – wpływ wielkości nacisku i czasu na wynik oznaczenia, Inżynieria Przetwórstwa Spożywczego, 1, 23-25.
7. Pisuła Andrzej., Pośpiech Edward., 2011, Mięso- podstawy nauki i technologii Pisula A., Pospiech E., 2011, Mięso – podstawy nauki i technologii, wyd. SGGW, Warszawa, 15-21.
8. Gońi S.M., Salvadori V.O., 2010, Prediction of cooking times and weight losses during meat roasting, Journal of Food Engineering, 100(1), 1-11.
9. Sman R.G.M., Voda A., Dalen G., Dujster A., 2013, Ice crystal interspacing in frozen foods, Journal of Food Engineering, 116 (2), 622-626.
10. Stodolnik L., Rosiński P., Grzegrzółka A., 2004, Krioochronne właściwości pektyny i lecytyny w tkance mięśniowej drobiu w czasie zamrażalniczego przechowywania, Chłodnictwo: organ Naczelnej Organizacjii Technicznej, 39(8), 74-77.
11. Tomkiewicz D. Seńcio M., Dolik K., 2012, Układ pomiarowy do określenia parametrów tekstury mięsa na podstawie pomiaru zmiany prędkości wycieku wody, Pomiary Automatyka Kontrola, vol 58 (10), 901-904.
12. Volpato G., Michielin E.M.Z., Ferreira S.R.S., Petrus J.C.C., 2008, Optimization of the chicken breast cooking process, Journal of Food Engineering, 84 (4), 576-581.
13. Barbera S., Tassone S., 2006, Meat cooking shrinkage: Measurement of a new meat quality parameter, Meat Science, 73, 467-474.
14. Prost E., 2006, Zwierzęta rzeźne i mięso – ocena i higiena. Lubelskie Towarzystwo Naukowe, Lublin
15. Pałacha Z., 2008, Aktywność wody ważny parametr trwałości żywności. Przemysł Spożywczy 4, 22-26.
16. Orkusz A., 2015, Czynniki kształtujące jakość mięsa drobiu grzebiącego. Nauki Inżynierskie i Technologie. Tom 1, Nr 16, 52-53.
17. Borzyszkowski M., 2015, Właściwości technologiczne mięsa wołowego, Wielka księga wołowiny; QMP-dobra wołowina, 103-106.
18. Domaradzki P., Litwińczuk Z., Florek M., Litwińczuk A., 2016, Zmiany właściwości fizykochemicznych i sensorycznych mięsa wołowego w zależności od warunków jego dojrzewania, Żywność. Nauka. Technologia. Jakość. Tom 23, Nr 3, 35-53 (Lesiów 2001).